|
|
|
A comprehensive analysis of immunoglobulin heavy chain genes in the Bactrian camel (Camelus bactrianus) |
Zuoxiang LIANG1,2,Tao WANG2,Yi SUN3,Wenlong YANG1,Zhihong LIU4,Jing FEI2,Ying GUO2,Qingwei MA5,Qingjie PAN1,*( ),Liming REN2,*() ),Liming REN2,*() |
1. Key Laboratory of Animal Reproduction and Germplasm Enhancement in Universities of Shandong, College of Animal Science and Technology, Qingdao Agricultural University, Qingdao 266109, China
2. State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing 100193, China
3. Laboratory of Animal Molecular Genetics, College of Animal Science and Veterinary Medicine, Shandong Agricultural University, Taian 271018, China
4. Key Laboratory of Animal Genetics, Breeding and Reproduction, Inner Mongolia Agricultural University, Hohhot 010018, China
5. Bioyong Technology Inc., Beijing 100080, China |
|
|
|
|
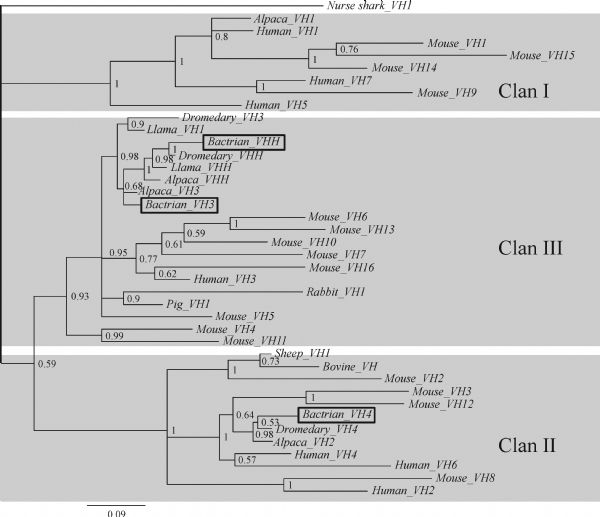
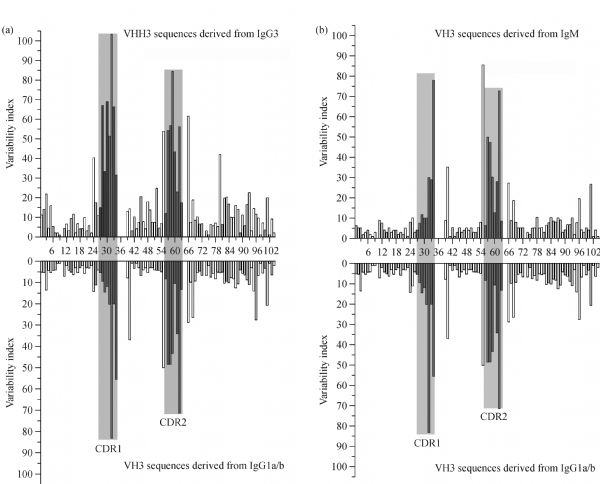
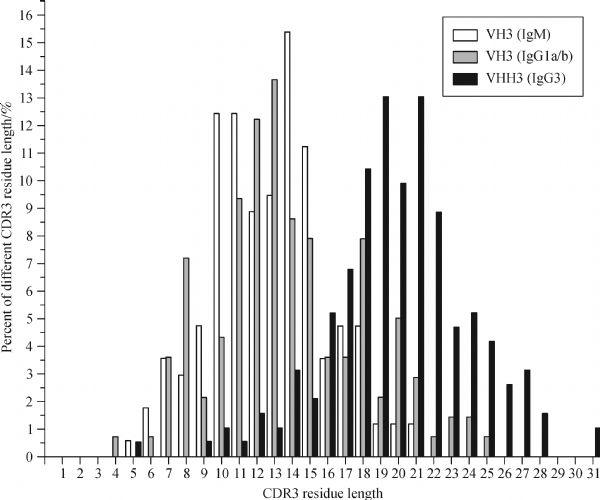
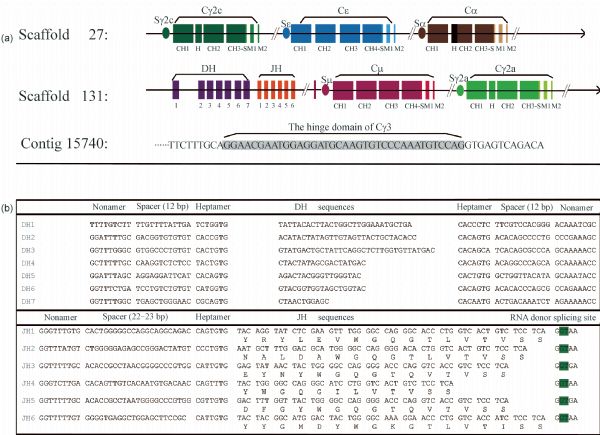
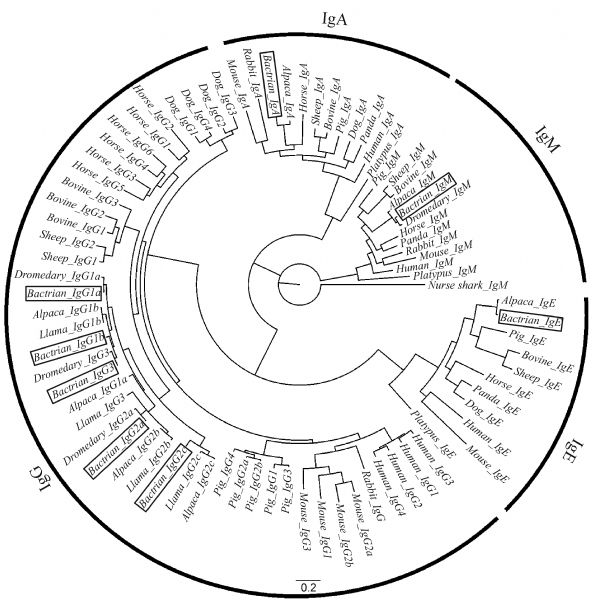
Abstract Heavy chain only antibodies (HCAbs) represent a rare type of antibody that is devoid of light chains and the CH1 domain that have been reported in cartilaginous fish and camelids. By analyzing transcript data and genome sequences, we conducted a comprehensive analysis of Bactrian camel immunoglobulin heavy chain genes. Based on the transcript data, one μ gene, five γ genes, one α gene and one ε gene were found. Additionally, the variable region of HCAbs (VHH) and the conventional antibodies (VH) sequences associated with the γ3, γ1a/b and μ genes were amplified. Based on these genome sequences, seven DH, six JH, μ, γ2a, γ2c, α, and ε genes and a portion of a γ3 gene were observed. Different Kozak sequences within different VH families were found in our analysis, and the variability index differed between the VHH3 and VH3 families. Phylogenetic analysis of the constant regions of the camelid immunoglobulin genes indicates that these genes appeared before the evolutionary divergence of Bactrian camels and dromedaries.
|
| Keywords
Bactrian camel
heavy-chain antibodies
VHH
γ3
|
|
Corresponding Author(s):
Qingjie PAN,Liming REN
|
|
Just Accepted Date: 28 May 2015
Online First Date: 18 June 2015
Issue Date: 10 November 2015
|
|
| 1 |
Edelman G M. Antibody structure and molecular immunology. Scandinavian Journal of Immunology, 1991, 34(1): 4−22
https://doi.org/10.1111/j.1365-3083.1991.tb01516.x
pmid: 2068529
|
| 2 |
Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hammers C, Songa E B, Bendahman N, Hammers R. Naturally occurring antibodies devoid of light chains. Nature, 1993, 363(6428): 446−448
https://doi.org/10.1038/363446a0
pmid: 8502296
|
| 3 |
Seligmann M, Mihaesco E, Preud’homme J L, Danon F, Brouet J C. Heavy chain diseases: current findings and concepts. Immunological Reviews, 1979, 48(1): 145−167
https://doi.org/10.1111/j.1600-065X.1979.tb00302.x
pmid: 121099
|
| 4 |
Dooley H, Flajnik M F. Antibody repertoire development in cartilaginous fish. Developmental and Comparative Immunology, 2006, 30(1−2): 43−56
https://doi.org/10.1016/j.dci.2005.06.022
pmid: 16146649
|
| 5 |
Flajnik M F, Deschacht N, Muyldermans S. A case of convergence: why did a simple alternative to canonical antibodies arise in sharks and camels? PLoS Biology, 2011, 9(8): e1001120
https://doi.org/10.1371/journal.pbio.1001120
pmid: 21829328
|
| 6 |
De Genst E, Saerens D, Muyldermans S, Conrath K. Antibody repertoire development in camelids. Developmental and Comparative Immunology, 2006, 30(1−2): 187−198
https://doi.org/10.1016/j.dci.2005.06.010
pmid: 16051357
|
| 7 |
Muyldermans S, Atarhouch T, Saldanha J, Barbosa J A, Hamers R. Sequence and structure of VH domain from naturally occurring camel heavy chain immunoglobulins lacking light chains. Protein Engineering, 1994, 7(9): 1129−1135
https://doi.org/10.1093/protein/7.9.1129
pmid: 7831284
|
| 8 |
Muyldermans S, Cambillau C, Wyns L. Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains. Trends in Biochemical Sciences, 2001, 26(4): 230−235
https://doi.org/10.1016/S0968-0004(01)01790-X
pmid: 11295555
|
| 9 |
Desmyter A, Transue T R, Ghahroudi M A, Dao Thi M H, Poortmans F, Hamers R, Muyldermans S, Wyns L. Crystal structure of a camel single-domain VH antibody fragment in complex with lysozyme. Nature Structural Biology, 1996, 3(9): 803−811
https://doi.org/10.1038/nsb0996-803
pmid: 8784355
|
| 10 |
Govaert J, Pellis M, Deschacht N, Vincke C, Conrath K, Muyldermans S, Saerens D. Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. The Journal of Biological Chemistry, 2012, 287(3): 1970−1979
https://doi.org/10.1074/jbc.M111.242818
pmid: 22128183
|
| 11 |
Khong Nguyen V, Hamers R, Wyns L, Muyldermans S. Loss of splice consensus signal is responsible for the removal of the entire C(H)1 domain of the functional camel IGG2A heavy-chain antibodies. Molecular Immunology, 1999, 36(8): 515−524
https://doi.org/10.1016/S0161-5890(99)00067-X
pmid: 10475606
|
| 12 |
Wu Y, Eigenbrot C, Liang W C, Stawicki S, Shia S, Fan B, Ganesan R, Lipari M T, Kirchhofer D. Structural insight into distinct mechanisms of protease inhibition by antibodies. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(50): 19784−19789
https://doi.org/10.1073/pnas.0708251104
pmid: 18077410
|
| 13 |
Lauwereys M, Arbabi Ghahroudi M, Desmyter A, Kinne J, Hölzer W, De Genst E, Wyns L, Muyldermans S. Potent enzyme inhibitors derived from dromedary heavy-chain antibodies. The EMBO Journal, 1998, 17(13): 3512−3520
https://doi.org/10.1093/emboj/17.13.3512
pmid: 9649422
|
| 14 |
Jirimutu W Z, Wang Z, Ding G, Chen G, Sun Y, Sun Z, Zhang H, Wang L, Hasi S, Zhang Y, Li J, Shi Y, Xu Z, He C, Yu S, Li S, Zhang W, Batmunkh M, Ts B, Narenbatu, Unierhu, Bat-Ireedui S, Gao H, Baysgalan B, Li Q, Jia Z, Turigenbayila, Subudenggerile, Narenmanduhu, Wang Z, Wang J, Pan L, Chen Y, Ganerdene Y, Dabxilt, Erdemt, Altansha, Altansukh, Liu T, Cao M, Aruuntsever, Bayart, Hosblig, He F, Zha-ti A, Zheng G, Qiu F, Sun Z, Zhao L, Zhao W, Liu B, Li C, Chen Y, Tang X, Guo C, Liu W, Ming L, Temuulen, Cui A, Li Y, Gao J, Li J, Wurentaodi, Niu S, Sun T, Zhai Z, Zhang M, Chen C, Baldan T, Bayaer T, Li Y, Meng H. Genome sequences of wild and domestic bactrian camels. Nature Communications, 2012, 3: 1202
https://doi.org/10.1038/ncomms2192
pmid: 23149746
|
| 15 |
Wu H, Guang X, Al-Fageeh M B, Cao J, Pan S, Zhou H, Zhang L, Abutarboush M H, Xing Y, Xie Z, Alshanqeeti A S, Zhang Y, Yao Q, Al-Shomrani B M, Zhang D, Li J, Manee M M, Yang Z, Yang L, Liu Y, Zhang J, Altammami M A, Wang S, Yu L, Zhang W, Liu S, Ba L, Liu C, Yang X, Meng F, Wang S, Li L, Li E, Li X, Wu K, Zhang S, Wang J, Yin Y, Yang H, Al-Swailem A M, Wang J. Camelid genomes reveal evolution and adaptation to desert environments. Nature Communications, 2014, 5: 5188
https://doi.org/10.1038/ncomms6188
pmid: 25333821
|
| 16 |
Griffin L M, Snowden J R, Lawson A D G, Wernery U, Kinne J, Baker T S. Analysis of heavy and light chain sequences of conventional camelid antibodies from Camelus dromedarius and Camelus bactrianus species. Journal of Immunological Methods, 2014, 405: 35−46
https://doi.org/10.1016/j.jim.2014.01.003
pmid: 24444705
|
| 17 |
Achour I, Cavelier P, Tichit M, Bouchier C, Lafaye P, Rougeon F. Tetrameric and homodimeric camelid IgGs originate from the same IgH locus. Journal of Immunology, 2008, 181(3): 2001−2009
https://doi.org/10.4049/jimmunol.181.3.2001
pmid: 18641337
|
| 18 |
Conrath K E, Wernery U, Muyldermans S, Nguyen V K. Emergence and evolution of functional heavy-chain antibodies in Camelidae. Developmental and Comparative Immunology, 2003, 27(2): 87−103
https://doi.org/10.1016/S0145-305X(02)00071-X
pmid: 12543123
|
| 19 |
Giudicelli V, Lefranc M P. Ontology for immunogenetics: the IMGT-ONTOLOGY. Bioinformatics, 1999, 15(12): 1047−1054
https://doi.org/10.1093/bioinformatics/15.12.1047
pmid: 10745995
|
| 20 |
Fu X, Gao X, He S, Huang D, Zhang P, Wang X, Zhang S, Dang R, Yin S, Du E, Yang Z. Design and selection of a camelid single-chain antibody yeast two-hybrid library produced de novo for the cap protein of porcine circovirus type 2 (PCV2). PLoS ONE, 2013, 8(3): e56222
https://doi.org/10.1371/journal.pone.0056222
pmid: 23469171
|
| 21 |
Deschacht N, De Groeve K, Vincke C, Raes G, De Baetselier P, Muyldermans S. A novel promiscuous class of camelid single-domain antibody contributes to the antigen-binding repertoire. Journal of Immunology, 2010, 184(10): 5696−5704
https://doi.org/10.4049/jimmunol.0903722
pmid: 20404276
|
| 22 |
Nguyen V K, Hamers R, Wyns L, Muyldermans S. Camel heavy-chain antibodies: diverse germline V(H)H and specific mechanisms enlarge the antigen-binding repertoire. The EMBO Journal, 2000, 19(5): 921−930
https://doi.org/10.1093/emboj/19.5.921
pmid: 10698934
|
| 23 |
Maass D R, Sepulveda J, Pernthaner A, Shoemaker C B. Alpaca (Lama pacos) as a convenient source of recombinant camelid heavy chain antibodies (VHHs). Journal of Immunological Methods, 2007, 324(1−2): 13−25
https://doi.org/10.1016/j.jim.2007.04.008
pmid: 17568607
|
| 24 |
Johansson J, Aveskogh M, Munday B, Hellman L. Heavy chain V region diversity in the duck-billed platypus (Ornithorhynchus anatinus): long and highly variable complementarity-determining region 3 compensates for limited germline diversity. Journal of Immunology, 2002, 168(10): 5155−5162
https://doi.org/10.4049/jimmunol.168.10.5155
pmid: 11994470
|
| 25 |
Stanfield R L, Dooley H, Flajnik M F, Wilson I A. Crystal structure of a shark single-domain antibody V region in complex with lysozyme. Science, 2004, 305(5691): 1770−1773
https://doi.org/10.1126/science.1101148
pmid: 15319492
|
| 26 |
Vu K B, Ghahroudi M A, Wyns L, Muyldermans S. Comparison of llama VH sequences from conventional and heavy chain antibodies. Molecular Immunology, 1997, 34(16−17): 1121−1131
https://doi.org/10.1016/S0161-5890(97)00146-6
pmid: 9566760
|
| 27 |
Sircar A, Sanni K A, Shi J, Gray J J. Analysis and modeling of the variable region of camelid single-domain antibodies. Journal of Immunology, 2011, 186(11): 6357−6367
https://doi.org/10.4049/jimmunol.1100116
pmid: 21525384
|
| 28 |
Crooks G E, Hon G, Chandonia J M, Brenner S E. WebLogo: a sequence logo generator. Genome Research, 2004, 14(6): 1188−1190
https://doi.org/10.1101/gr.849004
pmid: 15173120
|
| 29 |
Sørensen V, Sundvold V, Michaelsen T E, Sandlie I. Polymerization of IgA and IgM: roles of Cys309/Cys414 and the secretory tailpiece. Journal of Immunology, 1999, 162(6): 3448−3455
pmid: 10092800
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|