|
|
|
METABOLIC AND TRANSCRIPTOME ANALYSIS REVEALS METABOLITE VARIATION AND FLAVONOID REGULATORY NETWORKS IN FRESH SHOOTS OF TEA (CAMELLIA SINENSIS) OVER THREE SEASONS |
Chen-Kai JIANG1,2, De-Jiang NI3, Ming-Zhe YAO1, Jian-Qiang MA1( ), Liang CHEN1() ), Liang CHEN1() |
1. Key Laboratory of Tea Biology and Resources Utilization, Ministry of Agriculture and Rural Affairs, Tea Research Institute of the Chinese Academy of Agricultural Sciences, Hangzhou 310008, China.
2. State Key Laboratory for Quality and Safety of Agro-products, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China.
3. College of Horticulture and Forestry Science, Huazhong Agricultural University, Wuhan 430070, China. |
|
|
|
|
Abstract • Metabolites of fresh tea shoots at harvest were profiled. • Season-dependent metabolites were identified. • Key genes responsible for flavonoid metabolism are proposed. • Regulated relationships among the main compounds were investigated.
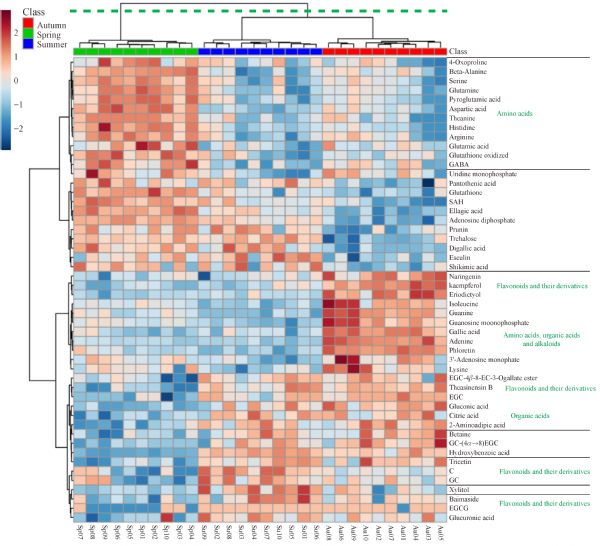
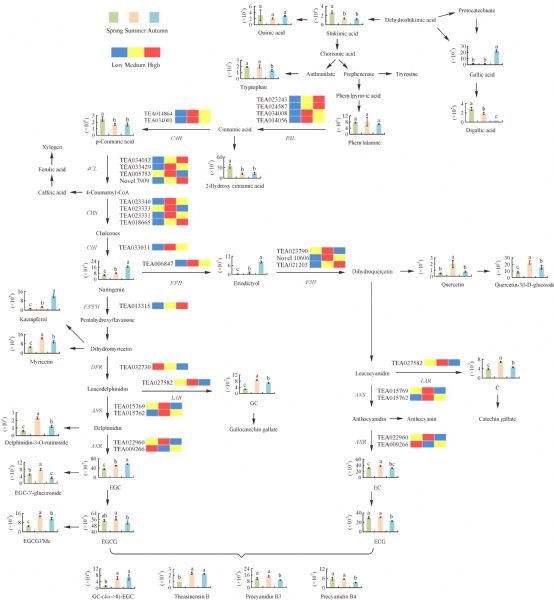
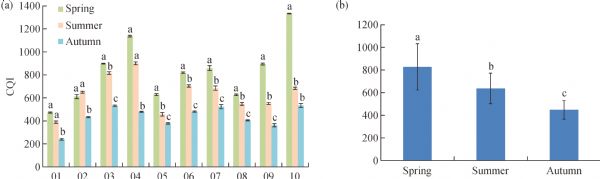
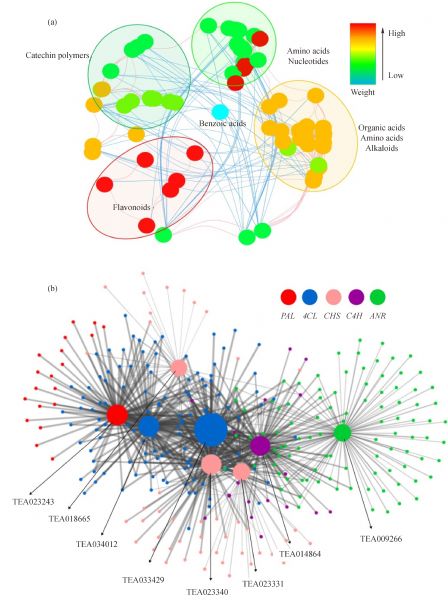
Metabolites, especially secondary metabolites, are very important in the adaption of tea plants and the quality of tea products. Here, we focus on the seasonal variation in metabolites of fresh tea shoots and their regulatory mechanism at the transcriptional level. The metabolic profiles of fresh tea shoots of 10 tea accessions collected in spring, summer, and autumn were analyzed using ultra-performance liquid chromatography coupled with quadrupole-obitrap mass spectrometry. We focused on the metabolites and key genes in the phenylpropanoid/flavonoid pathway integrated with transcriptome analysis. Multivariate statistical analysis indicates that metabolites were distinctly different with seasonal alternation. Flavonoids, amino acids, organic acids and alkaloids were the predominant metabolites. Levels of most key genes and downstream compounds in the flavonoid pathway were lowest in spring but the catechin quality index was highest in spring. The regulatory pathway was explored by constructing a metabolite correlation network and a weighted gene co-expression network.
|
| Keywords
harvest season
metabolites
tea shoots
transcriptomics
untargeted metabolomics
|
|
Corresponding Author(s):
Jian-Qiang MA,Liang CHEN
|
|
Just Accepted Date: 20 January 2021
Online First Date: 02 March 2021
Issue Date: 13 July 2021
|
|
| 1 |
N Sanlier, B B Gokcen, M Altuğ. Tea consumption and disease correlations. Trends in Food Science & Technology, 2018, 78: 95–106
https://doi.org/10.1016/j.tifs.2018.05.026
|
| 2 |
H G Ji, Y R Lee, M S Lee, K H Hwang, E H Kim, J S Park, Y S Hong. Metabolic phenotyping of various tea (Camellia sinensis L.) cultivars and understanding of their intrinsic metabolism. Food Chemistry, 2017, 233: 321–330
https://doi.org/10.1016/j.foodchem.2017.04.079
pmid: 28530581
|
| 3 |
R Fang, S P Redfern, D Kirkup, E A Porter, G C Kite, L A Terry, M J Berry, M S J Simmonds. Variation of theanine, phenolic, and methylxanthine compounds in 21 cultivars of Camellia sinensis harvested in different seasons. Food Chemistry, 2017, 220: 517–526
https://doi.org/10.1016/j.foodchem.2016.09.047
pmid: 27855934
|
| 4 |
H Li, Z W Liu, Z J Wu, Y X Wang, R M Teng, J Zhuang. Differentially expressed protein and gene analysis revealed the effects of temperature on changes in ascorbic acid metabolism in harvested tea leaves. Horticulture Research, 2018, 5(1): 65
https://doi.org/10.1038/s41438-018-0070-x
pmid: 30302261
|
| 5 |
S Jayasekera, L Kaur, A L Molan, M L Garg, P J Moughan. Effects of season and plantation on phenolic content of unfermented and fermented Sri Lankan tea. Food Chemistry, 2014, 152: 546–551
https://doi.org/10.1016/j.foodchem.2013.12.005
pmid: 24444973
|
| 6 |
W Dai, D Qi, T Yang, H Lv, L Guo, Y Zhang, Y Zhu, Q Peng, D Xie, J Tan, Z Lin. Nontargeted analysis using ultraperformance liquid chromatography-quadrupole time-of-flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality of tea (Camellia sinensis L.). Journal of Agricultural and Food Chemistry, 2015, 63(44): 9869–9878
https://doi.org/10.1021/acs.jafc.5b03967
pmid: 26494158
|
| 7 |
P Zhou, O Hu, H Fu, L Ouyang, X Gong, P Meng, Z Wang, M Dai, X Guo, Y Wang. UPLC-Q-TOF/MS-based untargeted metabolomics coupled with chemometrics approach for Tieguanyin tea with seasonal and year variations. Food Chemistry, 2019, 283: 73–82
https://doi.org/10.1016/j.foodchem.2019.01.050
pmid: 30722928
|
| 8 |
Q Zhang, M Liu, J Ruan. Metabolomics analysis reveals the metabolic and functional roles of flavonoids in light-sensitive tea leaves. BMC Plant Biology, 2017, 17(1): 64
https://doi.org/10.1186/s12870-017-1012-8
pmid: 28320327
|
| 9 |
J Zhu, Q Xu, S Zhao, X Xia, X Yan, Y An, X Mi, L Guo, L Samarina, C Wei. Comprehensive co-expression analysis provides novel insights into temporal variation of flavonoids in fresh leaves of the tea plant (Camellia sinensis). Plant Science, 2020, 290: 110306
pmid: 31779914
|
| 10 |
J Xu, Q F Zhang, J Zheng, B F Yuan, Y Q Feng. Mass spectrometry-based fecal metabolome analysis. Trends in Analytical Chemistry, 2019, 112: 161–174
|
| 11 |
W Xu, Q Song, D Li, X Wan. Discrimination of the production season of Chinese green tea by chemical analysis in combination with supervised pattern recognition. Journal of Agricultural and Food Chemistry, 2012, 60(28): 7064–7070
pmid: 22720840
|
| 12 |
D Qi, J Li, X Qiao, M Lu, W Chen, A Miao, W Guo, C Ma. Non-targeted metabolomic analysis based on ultra-high-performance liquid chromatography quadrupole time-of-flight tandem mass spectrometry reveals the effects of grafting on non-volatile metabolites in fresh tea leaves (Camellia sinensis L.). Journal of Agricultural and Food Chemistry, 2019, 67(23): 6672–6682
pmid: 31117493
|
| 13 |
X Guo, C T Ho, W Schwab, C Song, X Wan. Aroma compositions of large-leaf yellow tea and potential effect of theanine on volatile formation in tea. Food Chemistry, 2019, 280: 73–82
https://doi.org/10.1016/j.foodchem.2018.12.066
pmid: 30642509
|
| 14 |
M Kito, H Kokura, J Izaki, K Sasaoka. Theanine, a precursor of the phloroglucinol nucleus of catechins in tea plants. Phytochemistry, 1968, 7(4): 599–603
https://doi.org/10.1016/S0031-9422(00)88234-5
|
| 15 |
D N Barua. Seasonal dormancy in tea (Camellia sinensis L.). Nature, 1969, 224(5218): 514
https://doi.org/10.1038/224514a0
|
| 16 |
C Yue, H Cao, X Hao, J Zeng, W Qian, Y Guo, N Ye, Y Yang, X Wang. Differential expression of gibberellin- and abscisic acid-related genes implies their roles in the bud activity-dormancy transition of tea plants. Plant Cell Reports, 2018, 37(3): 425–441
https://doi.org/10.1007/s00299-017-2238-5
pmid: 29214380
|
| 17 |
T Tohge, L P de Souza, A R Fernie. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. Journal of Experimental Botany, 2017, 68(15): 4013–4028
https://doi.org/10.1093/jxb/erx177
pmid: 28922752
|
| 18 |
S Kaneko, K Kumazawa, H Masuda, A Henze, T Hofmann. Molecular and sensory studies on the umami taste of Japanese green tea. Journal of Agricultural and Food Chemistry, 2006, 54(7): 2688–2694
https://doi.org/10.1021/jf0525232
pmid: 16569062
|
| 19 |
Q Q Cao, C Zou, Y H Zhang, Q Z Du, J F Yin, J Shi, S Xue, Y Q Xu. Improving the taste of autumn green tea with tannase. Food Chemistry, 2019, 277: 432–437
https://doi.org/10.1016/j.foodchem.2018.10.146
pmid: 30502167
|
| 20 |
Y S Zhen, Z M Chen, S J Cheng, M L Chen. Tea: Bioactivity and Therapeutic Potential. London, UK: CRC Press, 2002
|
| 21 |
S Kallithraka, J Bakker, M N Clifford. Evaluation of bitterness and astringency of (+)-catechin and (–)-epicatechin in red wine and in model solution. Journal of Sensory Studies, 1997, 12(1): 25–37
https://doi.org/10.1111/j.1745-459X.1997.tb00051.x
|
| 22 |
H Peleg, K Gacon, P Schlich, A C Noble. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. Journal of the Science of Food and Agriculture, 1999, 79(8): 1123–1128
https://doi.org/10.1002/(SICI)1097-0010(199906)79:8<1123::AID-JSFA336>3.0.CO;2-D
|
| 23 |
C Liu, X Wang, V Shulaev, R A Dixon. A role for leucoanthocyanidin reductase in the extension of proanthocyanidins. Nature Plants, 2016, 2(12): 16182
https://doi.org/10.1038/nplants.2016.182
pmid: 27869786
|
| 24 |
R Stracke, H Ishihara, G Huep, A Barsch, F Mehrtens, K Niehaus, B Weisshaar. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant Journal, 2007, 50(4): 660–677
https://doi.org/10.1111/j.1365-313X.2007.03078.x
pmid: 17419845
|
| 25 |
S Chen, F Wu, Y Li, Y Qian, X Pan, F Li, Y Wang, Z Wu, C Fu, H Lin, A Yang. NtMYB4 and NtCHS1 are critical factors in the regulation of flavonoid biosynthesis and are involved in salinity responsiveness. Frontiers in Plant Science, 2019, 10: 178
https://doi.org/10.3389/fpls.2019.00178
pmid: 30846995
|
| 26 |
J I Kim, X Zhang, P E Pascuzzi, C J Liu, C Chapple. Glucosinolate and phenylpropanoid biosynthesis are linked by proteasome-dependent degradation of PAL. New Phytologist, 2020, 225(1): 154–168
https://doi.org/10.1111/nph.16108
pmid: 31408530
|
| 27 |
C Pastore, S Dal Santo, S Zenoni, N Movahed, G Allegro, G Valentini, I Filippetti, G B Tornielli. Whole plant temperature manipulation affects flavonoid metabolism and the transcriptome of grapevine berries. Frontiers in Plant Science, 2017, 8: 929
https://doi.org/10.3389/fpls.2017.00929
pmid: 28634482
|
| 28 |
Y Lu, Y Liu, X Niu, Q Yang, X Hu, H Y Zhang, J Xia. Systems genetic validation of the SNP-metabolite association in rice via metabolite-pathway-based phenome-wide association scans. Frontiers in Plant Science, 2015, 6: 1027
https://doi.org/10.3389/fpls.2015.01027
pmid: 26640468
|
| 29 |
Y C Ruan, Q K Cheng. The relation between the components of tea catechins and the quality of green tea. Acta Horticulturae Sinica, 1964, 3: 287–300 (in Chinese)
|
| 30 |
Y R Liang, J L Lu, L Y Zhang. Comparative study of cream in infusions of black tea and green tea (Camellia sinensis (L.) O. Kuntze). International Journal of Food Science & Technology, 2002, 37(6): 627–634
https://doi.org/10.1046/j.1365-2621.2002.00589.x
|
| 31 |
G A A R Perera, A M T Amarakoon, D C K Illeperuma, P K P Muthukumarana. Effects of raw material on the chemical composition, organoleptic properties, antioxidant activity, physical properties and the yield of instant black tea. Lebensmittel-Wissenschaft+Technologie, 2015, 63(1): 745–750
https://doi.org/10.1016/j.lwt.2015.03.060
|
| 32 |
B S Moore, C Hertweck, J N Hopke, M Izumikawa, J A Kalaitzis, G Nilsen, T O’Hare, J Piel, P R Shipley, L Xiang, M B Austin, J P Noel. Plant-like biosynthetic pathways in bacteria: from benzoic acid to chalcone. Journal of Natural Products, 2002, 65(12): 1956–1962
https://doi.org/10.1021/np020230m
pmid: 12502351
|
| 33 |
C Zheng, J Q Ma, J D Chen, C L Ma, W Chen, M Z Yao, L Chen. Gene co-expression networks reveal key drivers of flavonoid variation in eleven tea cultivars (Camellia sinensis). Journal of Agricultural and Food Chemistry, 2019, 67(35): 9967–9978
https://doi.org/10.1021/acs.jafc.9b04422
pmid: 31403784
|
| 34 |
M Nemesio-Gorriz, P B Blair, K Dalman, A Hammerbacher, J Arnerup, J Stenlid, S M Mukhtar, M Elfstrand. Identification of norway spruce MYB-bHLH-WDR transcription factor complex members linked to regulation of the flavonoid pathway. Frontiers in Plant Science, 2017, 8: 305
https://doi.org/10.3389/fpls.2017.00305
pmid: 28337212
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|