|
|
|
UNREDUCED MEGAGAMETOPHYTE FORMATION VIA SECOND DIVISION RESTITUTION CONTRIBUTES TO TETRAPLOID PRODUCTION IN INTERPLOIDY CROSSES WITH ‘ORAH’ MANDARIN (CITRUS RETICULATA) |
Qiangming XIA1, Wei WANG1, Kaidong XIE1( ), Xiaomeng WU1, Xiuxin DENG1, Jude W. GROSSER2, Wenwu GUO1() ), Xiaomeng WU1, Xiuxin DENG1, Jude W. GROSSER2, Wenwu GUO1() |
1. Key Laboratory of Horticultural Plant Biology (Ministry of Education), College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Wuhan 430070, China.
2. Citrus Research and Education Center, University of Florida/IFAS, Lake Alfred, USA. |
|
|
|
|
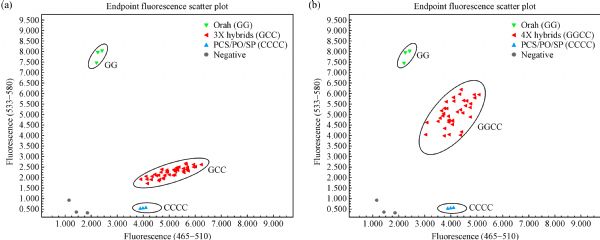
Abstract • In addition to triploid progeny, tetraploid hybrids derived from the fertilization of 2n megagametophytes are frequently regenerated from 2x × 4x crosses that utilize ‘Orah’ mandarin as the female parent. • Data here indicate that ‘Orah’ mandarin is a cultivar that readily produces 2n megagametophytes. • Second division restitution is the mechanism underlying 2n megagametophyte formation in ‘Orah’ mandarin.
Seedless fruits are desirable in the citrus fresh fruit market. Triploid production via diploid × tetraploid interploidy crosses is thought to be the most efficient and widely-used strategy for the breeding of seedless citrus. Although ‘Orah’ mandarin has desirable organoleptic qualities, seeds in the fruits weaken its market competitiveness. To produce new seedless cultivars that are similar to ‘Orah’ mandarin, we performed three 2x × 4x crosses using ‘Orah’ mandarin as the seed parent to regenerate triploid plantlets. A total of 182 triploid and 36 tetraploid plantlets were obtained. By analyzing their genetic origins using nine novel single nucleotide polymorphism (SNP) markers, all of the triploids and tetraploids derived from these three crosses were proven to be hybrids. Also, we demonstrated that 2n megagametophyte formation in ‘Orah’ mandarin result in tetraploid production in these three interploidy crosses. These tetraploid plantlets were genotyped using eight pericentromeric SNP markers and nine centromere distal SNP markers. Based on the genotypes of the 2n megagametophytes, the parental heterozygosity rates in 16 SNP loci and all 2n megagametophytes were less than 50%, indicating that second division restitution was the mechanism underlying 2n megagametophyte formation at both the population and individual levels. These triploid hybrids enrich the germplasm available for seedless breeding. Moreover, the tetraploid hybrids are valuable as parents for ploidy breeding for the production of seedless citrus fruits.
|
| Keywords
Citrus
2n gamete
interploidy hybridization
pericentromeric SNP marker
second division restitution
|
|
Corresponding Author(s):
Kaidong XIE,Wenwu GUO
|
|
Just Accepted Date: 03 February 2021
Online First Date: 15 March 2021
Issue Date: 13 July 2021
|
|
| 1 |
L Brownfield, C Köhler. Unreduced gamete formation in plants: mechanisms and prospects. Journal of Experimental Botany, 2011, 62(5): 1659–1668
https://doi.org/10.1093/jxb/erq371
pmid: 21109579
|
| 2 |
C A Krug. Chromosome numbers in the subfamily Arantioideae, with special reference in the genus Citrus. Botanical Gazette, 1943, 104(4): 602–611
https://doi.org/10.1086/335173
|
| 3 |
P Aleza, J Cuenca, J Juárez, L Navarro, P Ollitrault. Inheritance in doubled-diploid clementine and comparative study with SDR unreduced gametes of diploid clementine. Plant Cell Reports, 2016, 35(8): 1573–1586
https://doi.org/10.1007/s00299-016-1972-4
pmid: 27038940
|
| 4 |
P Ollitrault, M A Germana, Y Froelicher, J Cuenca, P Aleza, R Morillon, J W Grosser, W W Guo. Ploidy Manipulation for citrus breeding, genetics, and genomics. In: Gentile A, La Malfa S, Deng Z, eds. The Citrus Genome. Springer, 2020, 75–105
|
| 5 |
P Ollitrault, D Dambier, F Luro, Y Froelicher. Ploidy manipulation for breeding seedless triploid citrus. In: Janick J, ed. Plant Breeding Reviews. Wiley, 2008, 30: 323–352
|
| 6 |
G R Recupero, G Russo, S Recupero. New promising Citrus triploid hybrids selected from crosses between monoembryonic diploid female and tetraploid male parents. HortScience, 2005, 40(3): 516–520
https://doi.org/10.21273/HORTSCI.40.3.516
|
| 7 |
P Aleza, J Juárez, J Cuenca, P Ollitrault, L Navarro. Extensive citrus triploid hybrid production by 2x × 4x sexual hybridizations and parent-effect on the length of the juvenile phase. Plant Cell Reports, 2012, 31(9): 1723–1735
https://doi.org/10.1007/s00299-012-1286-0
pmid: 22614256
|
| 8 |
K D Xie, H Q Wang, X P Wang, W J Liang, Z Z Xie, H L Yi, X X Deng, J W Grosser, W W Guo. Extensive citrus triploid breeding by crossing monoembryonic diploid females with allotetraploid male parents. Scientia Agricultura Sinica, 2013, 46(21): 4550–4557 (in Chinese)
|
| 9 |
K D Xie, X P Wang, H Q Wang, W J Liang, Z Z Xie, D Y Guo, H L Yi, X X Deng, J W Grosser, W W Guo. High efficient and extensive production of triploid Citrus plants by crossing polyembryonic diploids with tetraploids. Acta Horticulturae Sinica, 2014, 41(4): 613–620 (in Chinese)
|
| 10 |
K D Xie, D Y Yuan, W Wang, Q M Xia, X M Wu, C W Chen, C L Chen, J W Grosser, W W Guo. Citrus triploid recovery based on 2x × 4x crosses via an optimized embryo rescue approach. Scientia Horticulturae, 2019, 252: 104–109
https://doi.org/10.1016/j.scienta.2019.03.038
|
| 11 |
J Cuenca, Y Froelicher, P Aleza, J Juárez, L Navarro, P Ollitrault. Multilocus half-tetrad analysis and centromere mapping in citrus: evidence of SDR mechanism for 2n megagametophyte production and partial chiasma interference in mandarin cv ‘Fortune’. Heredity, 2011, 107(5): 462–470
https://doi.org/10.1038/hdy.2011.33
pmid: 21587302
|
| 12 |
J M Kreiner, P Kron, B C Husband. Frequency and maintenance of unreduced gametes in natural plant populations: associations with reproductive mode, life history and genome size. New Phytologist, 2017, 214(2): 879–889
https://doi.org/10.1111/nph.14423
pmid: 28134436
|
| 13 |
P Aleza, J Cuenca, M Hernández, J Juárez, L Navarro, P Ollitrault. Genetic mapping of centromeres in the nine Citrus clementina chromosomes using half-tetrad analysis and recombination patterns in unreduced and haploid gametes. BMC Plant Biology, 2015, 15(1): 80–93
https://doi.org/10.1186/s12870-015-0464-y
pmid: 25848689
|
| 14 |
J Cuenca, P Aleza, J Juárez, A García-Lor, Y Froelicher, L Navarro, P Ollitrault. Maximum-likelihood method identifies meiotic restitution mechanism from heterozygosity transmission of centromeric loci: application in citrus. Scientific Reports, 2015, 5(1): 9897–9908
https://doi.org/10.1038/srep09897
pmid: 25894579
|
| 15 |
K D Xie, X P Wang, M K Biswas, W J Liang, Q Xu, J W Grosser, W W Guo. 2n megagametophyte formed via SDR contributes to tetraploidization in polyembryonic ‘Nadorcott’ tangor crossed by citrus allotetraploids. Plant Cell Reports, 2014, 33(10): 1641–1650
https://doi.org/10.1007/s00299-014-1643-2
pmid: 24972825
|
| 16 |
H Rouiss, J Cuenca, L Navarro, P Ollitrault, P Aleza. Tetraploid citrus progenies arising from FDR and SDR unreduced pollen in 4x × 2x hybridizations. Tree Genetics & Genomes, 2017, 13(1): 10–24
https://doi.org/10.1007/s11295-016-1094-8
|
| 17 |
H Rouiss, J Cuenca, L Navarro, P Ollitrault, P Aleza. Unreduced megagametophyte production in lemon occurs via three meiotic mechanisms, predominantly second-division restitution. Frontiers in Plant Science, 2017, 8: 1211–1227
https://doi.org/10.3389/fpls.2017.01211
pmid: 28747921
|
| 18 |
K D Xie, Q M Xia, J Peng, X M Wu, Z Z Xie, C L Chen, W W Guo. Mechanism underlying 2n male and female gamete formation in lemon via cytological and molecular marker analysis. Plant Biotechnology Reports, 2019, 13(2): 141–149
https://doi.org/10.1007/s11816-019-00525-4
|
| 19 |
G H Barry, F G Gmitter Jr, C X Chen, M L Roose, C T Federici, G T McCollum. Investigating the parentage of ‘Orri’ and ‘Fortune’ mandarin hybrids. Acta Horticulturae, 2015, (1065): 449–456
https://doi.org/10.17660/ActaHortic.2015.1065.55
|
| 20 |
W W Guo, D Prasad, P Serrano, F G Gmitter Jr, J W Grosser. Citrus somatic hybridization with potential for direct tetraploid scion cultivar development. Journal of Horticultural Science & Biotechnology, 2004, 79(3): 400–405
https://doi.org/10.1080/14620316.2004.11511780
|
| 21 |
J W Grosser, F G Gmitter Jr. Protoplast fusion for production of tetraploids and triploids: applications for scion and rootstock breeding in citrus. Plant Cell, Tissue and Organ Culture, 2011, 104(3): 343–357
https://doi.org/10.1007/s11240-010-9823-4
|
| 22 |
W W Guo, R C Wu, Y J Cheng, X X Deng. Production and molecular characterization of Citrus intergeneric somatic hybrids between red tangerine and citrange. Plant Breeding, 2007, 126(1): 72–76
https://doi.org/10.1111/j.1439-0523.2006.01315.x
|
| 23 |
S M Wang, H Lan, H H Jia, K D Xie, X M Wu, C L Chen, W W Guo. Induction of parthenogenetic haploid plants using gamma irradiated pollens in ‘Hirado Buntan’ pummelo (Citrus grandis [L.] Osbeck). Scientia Horticulturae, 2016, 207: 233–239
https://doi.org/10.1016/j.scienta.2016.05.028
|
| 24 |
Y J Cheng, W W Guo, H L Yi, X M Pang, X X Deng. An efficient protocol for genomic DNA extraction from Citrus species. Plant Molecular Biology Reporter, 2003, 21(2): 177–178
https://doi.org/10.1007/BF02774246
|
| 25 |
Q M Xia, L K Miao, K D Xie, Z P Yin, X M Wu, C L Chen, J W Grosser, W W Guo. Localization and characterization of Citrus centromeres by combining half-tetrad analysis and CenH3-associated sequence profiling. Plant Cell Reports, 2020, 39(12): 1609–1622
https://doi.org/10.1007/s00299-020-02587-z
pmid: 32897396
|
| 26 |
Q Xu, L L Chen, X Ruan, D Chen, A Zhu, C Chen, D Bertrand, W B Jiao, B H Hao, M P Lyon, J Chen, S Gao, F Xing, H Lan, J W Chang, X Ge, Y Lei, Q Hu, Y Miao, L Wang, S Xiao, M K Biswas, W Zeng, F Guo, H Cao, X Yang, X W Xu, Y J Cheng, J Xu, J H Liu, O J Luo, Z Tang, W W Guo, H Kuang, H Y Zhang, M L Roose, N Nagarajan, X X Deng, Y Ruan. The draft genome of sweet orange (Citrus sinensis). Nature Genetics, 2013, 45(1): 59–66
https://doi.org/10.1038/ng.2472
pmid: 23179022
|
| 27 |
H Li, R Durbin. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics, 2010, 26(5): 589–595
https://doi.org/10.1093/bioinformatics/btp698
pmid: 20080505
|
| 28 |
H Li, B Handsaker, A Wysoker, T Fennell, J Ruan, N Homer, G Marth, G Abecasis, R Durbin. The Sequence Alignment/Map format and SAMtools. Bioinformatics, 2009, 25(16): 2078–2079
https://doi.org/10.1093/bioinformatics/btp352
pmid: 19505943
|
| 29 |
P Cingolani, A Platts, L Wang, M Coon, T Nguyen, L Wang, S J Land, X Lu, D M Ruden. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly, 2012, 6(2): 80–92
https://doi.org/10.4161/fly.19695
pmid: 22728672
|
| 30 |
J Cuenca, P Aleza, L Navarro, P Ollitrault. Assignment of SNP allelic configuration in polyploids using competitive allele-specific PCR: application to citrus triploid progeny. Annual of Botany, 2013, 111(4): 731–742
https://doi.org/10.1093/aob/mct032
pmid: 23422023
|
| 31 |
A Blary, E Jenczewski. Manipulation of crossover frequency and distribution for plant breeding. Theoretical and Applied Genetics, 2019, 132(3): 575–592
https://doi.org/10.1007/s00122-018-3240-1
pmid: 30483818
|
| 32 |
J B Fernandes, P Wlodzimierz, I R Henderson. Meiotic recombination within plant centromeres. Current Opinion in Plant Biology, 2019, 48: 26–35
https://doi.org/10.1016/j.pbi.2019.02.008
pmid: 30954771
|
| 33 |
C B Dong, Y J Suo, X Y Kang. Assessment of the genetic composition of triploid hybrid Populus using SSR markers with low recombination frequencies. Canadian Journal of Forest Research, 2014, 44(7): 692–699
https://doi.org/10.1139/cjfr-2013-0360
|
| 34 |
H Liesebach, K Ulrich, D Ewald. FDR and SDR processes in meiosis and diploid gamete formation in poplars (Populus L.) detected by centromere-associated microsatellite markers. Tree Genetics & Genomes, 2015, 11(1): 801–811
https://doi.org/10.1007/s11295-014-0801-6
|
| 35 |
J Cuenca, P Aleza, A Garcia-Lor, P Ollitrault, L Navarro. Fine mapping for identification of Citrus alternaria brown spot candidate resistance genes and development of new SNP markers for marker-assisted selection. Frontiers in Plant Science, 2016, 7: 1948–1961
https://doi.org/10.3389/fpls.2016.01948
pmid: 28066498
|
| 36 |
X Wang, Y Xu, S Zhang, L Cao, Y Huang, J Cheng, G Wu, S Tian, C Chen, Y Liu, H Yu, X Yang, H Lan, N Wang, L Wang, J Xu, X Jiang, Z Xie, M Tan, R M Larkin, L L Chen, B G Ma, Y Ruan, X Deng, Q Xu. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nature Genetics, 2017, 49(5): 765–772
https://doi.org/10.1038/ng.3839
pmid: 28394353
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|