|
|
|
Engineering of β-carotene hydroxylase and ketolase for astaxanthin overproduction in Saccharomyces cerevisiae |
Ruizhao Wang1,2,Xiaoli Gu1,2,Mingdong Yao1,2,Caihui Pan1,2,Hong Liu1,2,Wenhai Xiao1,2( ),Ying Wang1,2(),Yingjin Yuan1,2 ),Ying Wang1,2(),Yingjin Yuan1,2 |
1. Key Laboratory of Systems Bioengineering (Ministry of Education), Tianjin University, Tianjin 300072, China
2. SynBio Research Platform, Collaborative Innovation Center of Chemical Science and Engineering (Tianjin), School of Chemical Engineering and Technology, Tianjin University, Tianjin 300072, China |
|
|
|
|
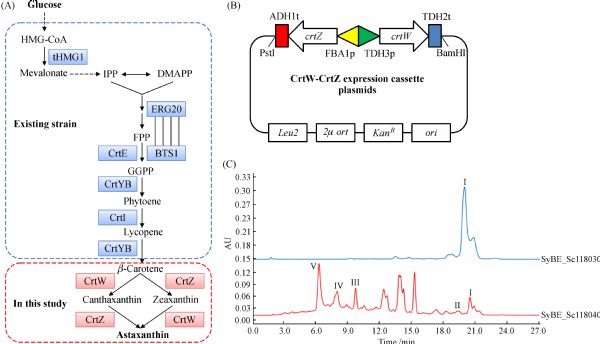
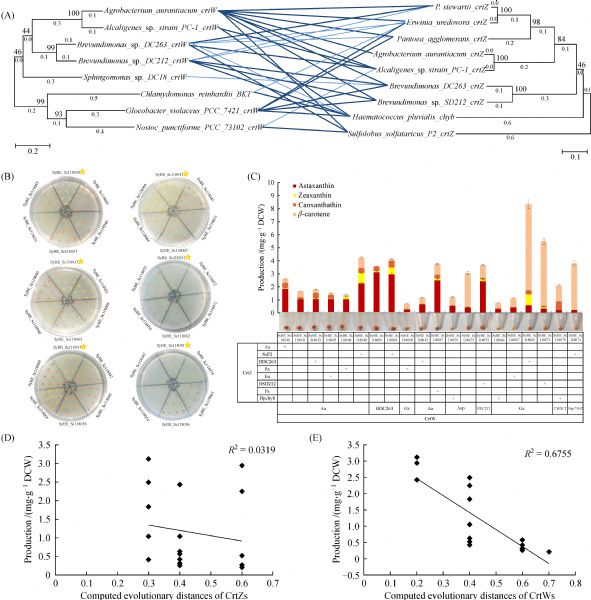
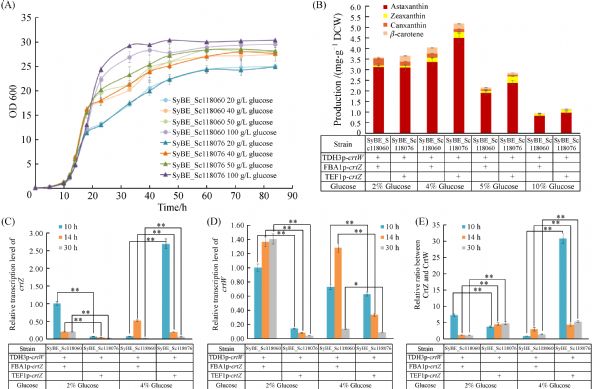
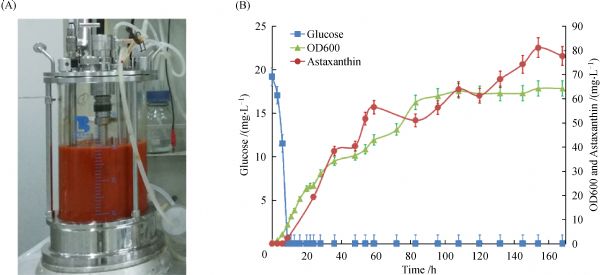
Abstract The conversion of β-carotene to astaxanthin is a complex pathway network, in which two steps of hydroxylation and two steps of ketolation are catalyzed by β-carotene hydroxylase (CrtZ) and β-carotene ketolase (CrtW) respectively. Here, astaxanthin biosynthesis pathway was constructed in Saccharomyces cerevisiae by introducing heterologous CrtZ and CrtW into an existing high β-carotene producing strain. Both genes crtZ and crtW were codon optimized and expressed under the control of constitutive promoters. Through combinatorial expression of CrtZ and CrtW from diverse species, nine strains in dark red were visually chosen from thirty combinations. In all the selected strains, strain SyBE_Sc118060 with CrtW from Brevundimonas vesicularis DC263 and CrtZ from Alcaligenes sp. strain PC-1 achieved the highest astaxanthin yield of 3.1 mg/g DCW. Protein phylogenetic analysis shows that the shorter evolutionary distance of CrtW is, the higher astaxanthin titer is. Further, when the promoter of crtZ in strain SyBE_Sc118060 was replaced from FBA1p to TEF1p, the astaxanthin yield was increased by 30.4% (from 3.4 to 4.5 mg/g DCW). In the meanwhile, 33.5-fold increase on crtZ transcription level and 39.1-fold enhancement on the transcriptional ratio of crtZ to crtW were observed at early exponential phase in medium with 4% (w/v) glucose. Otherwise, although the ratio of crtZ to crtW were increased at mid-, late-exponential phases in medium with 2% (w/v) glucose, the transcription level of both crtZ and crtW were actually decreased during the whole time course, consequently leading to no significant improvement on astaxanthin production. Finally, through high cell density fed-batch fermentation using a carbon source restriction strategy, the production of astaxanthin in a 5-L bioreactor reached to 81.0 mg/L, which was the highest astaxanthin titer reported in yeast. This study provides a reference to greatly enhance desired compounds accumulation by employing the key enzyme(s) in microbes.
|
| Keywords
synthetic biology
astaxanthin
β-carotene hydroxylase
β-carotene ketolase
Saccharomyces cerevisiae
|
|
|
| Fund: |
|
Corresponding Author(s):
Wenhai Xiao,Ying Wang
|
|
Just Accepted Date: 23 January 2017
Online First Date: 24 February 2017
Issue Date: 17 March 2017
|
|
| 1 |
Ambati R R, Phang S M, Ravi S, Aswathanarayana R G. Astaxanthin: sources, extraction, stability, biological activities and its commercial applications—a review. Marine Drugs, 2014, 12(1): 128–152
https://doi.org/10.3390/md12010128
|
| 2 |
Zhou P, Ye L, Xie W, Lv X, Yu H. Highly efficient biosynthesis of astaxanthin in Saccharomyces cerevisiae by integration and tuning of algal crtZ and bkt. Applied Microbiology and Biotechnology, 2015, 99(20): 8419–8428
https://doi.org/10.1007/s00253-015-6791-y
|
| 3 |
Ukibe K, Hashida K, Yoshida N, Takagi H. Metabolic engineering of Saccharomyces cerevisiae for astaxanthin production and oxidative stress tolerance. Applied and Environmental Microbiology, 2009, 75(22): 7205–7211
https://doi.org/10.1128/AEM.01249-09
|
| 4 |
Martin J F, Gudina E, Barredo J L. Conversion of beta-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein. Microbial Cell Factories, 2008, 7(1): 3
https://doi.org/10.1186/1475-2859-7-3
|
| 5 |
Chang J J, Thia C, Lin H Y, Liu H L, Ho F J, Wu J T, Shih M C, Li W H, Huang C C. Integrating an algal beta-carotene hydroxylase gene into a designed carotenoid-biosynthesis pathway increases carotenoid production in yeast. Bioresource Technology, 2015, 184: 2–8
https://doi.org/10.1016/j.biortech.2014.11.097
|
| 6 |
Sarria S, Wong B, Garcia M H, Keasling J D, Peralta-Yahya P. Microbial synthesis of pinene. ACS Synthetic Biology, 2014, 3(7): 466–475
https://doi.org/10.1021/sb4001382
|
| 7 |
Chen Y, Xiao W, Wang Y, Liu H, Li X, Yuan Y. Lycopene overproduction in Saccharomyces cerevisiae through combining pathway engineering with host engineering. Microbial Cell Factories, 2016, 15(1): 113
https://doi.org/10.1186/s12934-016-0509-4
|
| 8 |
Du H X, Xiao W H, Wang Y, Zhou X, Zhang Y, Liu D, Yuan Y J. Engineering Yarrowia lipolytica for campesterol overproduction. PLoS One, 2016, 11(1): e0146773
https://doi.org/10.1371/journal.pone.0146773
|
| 9 |
Fraser P D, Miura Y, Misawa N. In vitro characterization of astaxanthin biosynthetic enzymes. Journal of Biological Chemistry, 1997, 272(10): 6128–6135
https://doi.org/10.1074/jbc.272.10.6128
|
| 10 |
Ye R W, Stead K J, Yao H, He H. Mutational and functional analysis of the beta-carotene ketolase involved in the production of canthaxanthin and astaxanthin. Applied and Environmental Microbiology, 2006, 72(9): 5829–5837
https://doi.org/10.1128/AEM.00918-06
|
| 11 |
Tao L, Wilczek J, Odom J M, Cheng Q. Engineering a beta-carotene ketolase for astaxanthin production. Metabolic Engineering, 2006, 8(6): 523–531
https://doi.org/10.1016/j.ymben.2006.06.001
|
| 12 |
Scaife M A, Burja A M, Wright P C. Characterization of cyanobacterial beta-carotene ketolase and hydroxylase genes in Escherichia coli, and their application for astaxanthin biosynthesis. Biotechnology and Bioengineering, 2009, 103(5): 944–955
https://doi.org/10.1002/bit.22330
|
| 13 |
Choi S K, Nishida Y, Matsuda S, Adachi K, Kasai H, Peng X, Komemushi S, Miki W, Misawa N. Characterization of beta-carotene ketolases, CrtW, from marine bacteria by complementation analysis in Escherichia coli. Marine Biotechnology (New York, N.Y.), 2005, 7(5): 515–522
https://doi.org/10.1007/s10126-004-5100-z
|
| 14 |
Fraser P D, Shimada H, Misawa N. Enzymic confirmation of reactions involved in routes to astaxanthin formation, elucidated using a direct substrate in vitro assay. European Journal of Biochemistry, 1998, 252(2): 229–236
https://doi.org/10.1046/j.1432-1327.1998.2520229.x
|
| 15 |
Zelcbuch L, Antonovsky N, Bar-Even A, Levin-Karp A, Barenholz U, Dayagi M, Liebermeister W, Flamholz A, Noor E, Amram S, Spanning high-dimensional expression space using ribosome-binding site combinatorics. Nucleic Acids Research, 2013, 41(9): e98
https://doi.org/10.1093/nar/gkt151
|
| 16 |
Ajikumar P K, Xiao W H, Tyo K E, Wang Y, Simeon F, Leonard E, Mucha O, Phon T H, Pfeifer B, Stephanopoulos G. Isoprenoid pathway optimization for taxol precursor overproduction in Escherichia coli. Science, 2010, 330(6000): 70–74
https://doi.org/10.1126/science.1191652
|
| 17 |
Cao Y X, Xiao W H, Liu D, Zhang J L, Ding M Z, Yuan Y J. Biosynthesis of odd-chain fatty alcohols in Escherichia coli. Metabolic Engineering, 2015, 29: 113–123
https://doi.org/10.1016/j.ymben.2015.03.005
|
| 18 |
Lemuth K, Steuer K, Albermann C. Engineering of a plasmid-free Escherichia coli strain for improved in vivo biosynthesis of astaxanthin. Microbial Cell Factories, 2011, 10(1): 29
https://doi.org/10.1186/1475-2859-10-29
|
| 19 |
Wang R Z, Pan C H, Wang Y, Xiao W H, Yuan Y J. Design and construction of high β-carotene producing Saccharomyces cerevisiae. Journal of Chinese Biotechnology, 2016, 36: 83–91
|
| 20 |
Gietz R D, Schiestl R H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nature Protocols, 2007, 2(1): 31–34
https://doi.org/10.1038/nprot.2007.13
|
| 21 |
Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution, 2016, 33(7): 1870–1874
https://doi.org/10.1093/molbev/msw054
|
| 22 |
Saitou N, Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 1987, 4: 406–425
|
| 23 |
Sanderson M J, Wojciechowski M F. Improved bootstrap confidence limits in large-scale phylogenies, with an example from neo-astragalus (Leguminosae). Systematic Biology, 2000, 49: 671–685
https://doi.org/10.1080/106351500750049761
|
| 24 |
Zuckerkandl E, Pauling L. Evolutionary divergence and convergence in proteins. In: Bryson V, Vogel H J, eds. Evolving Genes and Proteins. New York: Academic Press, 1965, 97–166
|
| 25 |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods (San Diego, Calif.), 2001, 25(4): 402–408
https://doi.org/10.1006/meth.2001.1262
|
| 26 |
Jin Z, Wong A, Foo J L, Ng J, Cao Y X, Chang M W, Yuan Y J. Engineering Saccharomyces cerevisiae to produce odd chain-length fatty alcohols. Biotechnology and Bioengineering, 2016, 113(4): 842–851
https://doi.org/10.1002/bit.25856
|
| 27 |
Lorenz R T, Cysewski G R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends in Biotechnology, 2000, 18(4): 160–167
https://doi.org/10.1016/S0167-7799(00)01433-5
|
| 28 |
Scaife M A, Ma C A, Ninlayarn T, Wright P C, Armenta R E. Comparative analysis of beta-carotene hydroxylase genes for astaxanthin biosynthesis. Journal of Natural Products, 2012, 75(6): 1117–1124
https://doi.org/10.1021/np300136t
|
| 29 |
Nam H, Lewis N E, Lerman J A, Lee D H, Chang R L, Kim D, Palsson B O. Network context and selection in the evolution to enzyme specificity. Science, 2012, 337(6098): 1101–1104
https://doi.org/10.1126/science.1216861
|
| 30 |
Sandmann G. Evolution of carotene desaturation: The complication of a simple pathway. Archives of Biochemistry and Biophysics, 2009, 483(2): 169–174
https://doi.org/10.1016/j.abb.2008.10.004
|
| 31 |
Schaub P, Yu Q, Gemmecker S, Poussin-Courmontagne P, Mailliot J, McEwen A G, Ghisla S, Al-Babili S, Cavarelli J, Beyer P. On the structure and function of the phytoene desaturase CrtI from Pantoea ananatis, a membrane-peripheral and FAD-dependent oxidase/isomerase. PLoS One, 2012, 7(6): e39550
https://doi.org/10.1371/journal.pone.0039550
|
| 32 |
Sun J, Shao Z, Zhao H, Nair N, Wen F, Xu J H, Zhao H. Cloning and characterization of a panel of constitutive promoters for applications in pathway engineering in Saccharomyces cerevisiae. Biotechnology and Bioengineering, 2012, 109(8): 2082–2092
https://doi.org/10.1002/bit.24481
|
| 33 |
Peng B, Williams T C, Henry M, Nielsen L K, Vickers C E. Controlling heterologous gene expression in yeast cell factories on different carbon substrates and across the diauxic shift: A comparison of yeast promoter activities. Microbial Cell Factories, 2015, 14(1): 91
https://doi.org/10.1186/s12934-015-0278-5
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|