|
|
|
Depletion of conventional mature B cells and compromised specific antibody response in bovine immunoglobulin μ heavy-chain transgenic mice |
Min ZHANG1,Xueqian CHENG1,Dan CHU1,Jingwen LIANG1,Yi SUN1,Li MA1,Beilei XU1,Min ZHENG2,Meili WANG2,Liming REN1,Xiaoxiang HU1,Qingyong MENG1,Ran ZHANG1,Ying GUO1,Yunping DAI1,Robert AITKEN3,Ning LI1,Yaofeng ZHAO1,4,*( ) ) |
1. State Key Laboratory of Agrobiotechnology, College of Biological Sciences, China Agricultural University, Beijing 100193, China
2. GeneProtein Biotech Ltd, Beijing 100193, China
3. School of Life Sciences, College of Medical, Veterinary and Life Sciences, University of Glasgow, Glasgow, UK
4. Key Laboratory of Animal Reproduction and Germplasm Enhancement in Universities of Shandong, College of Animal Science and Technology, Qingdao Agricultural University, Qingdao 266109, China |
|
|
|
|
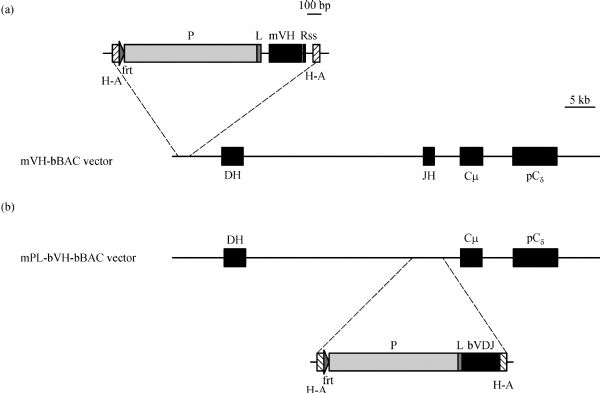
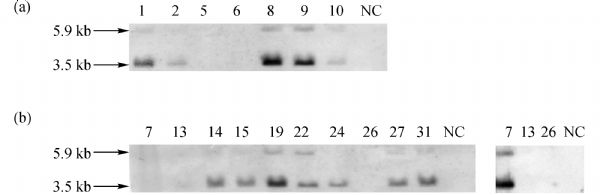
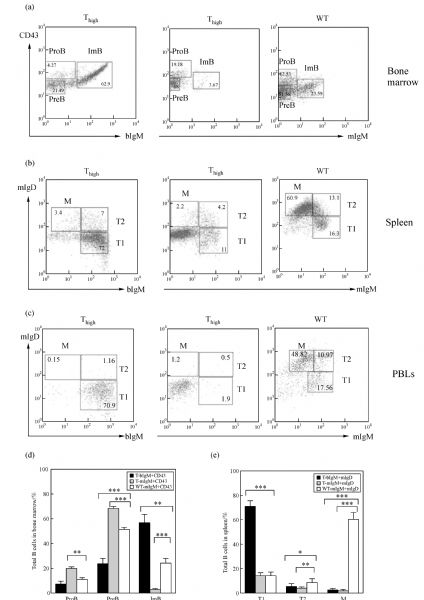
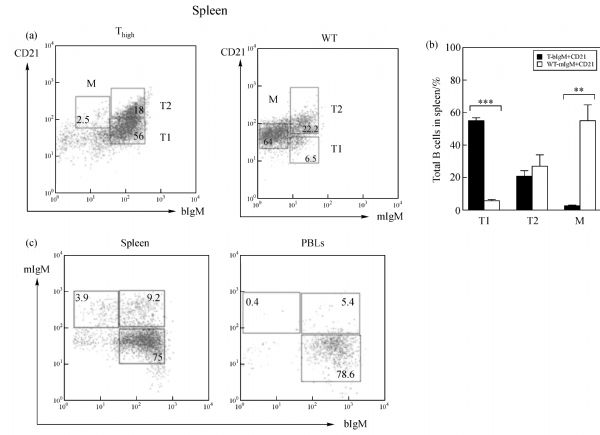
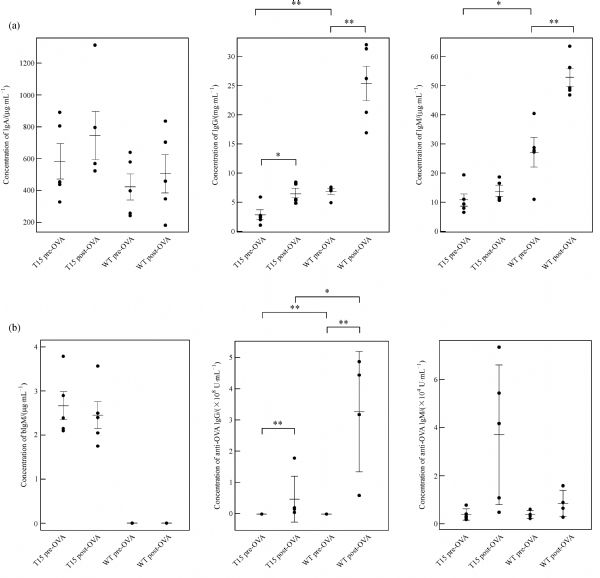
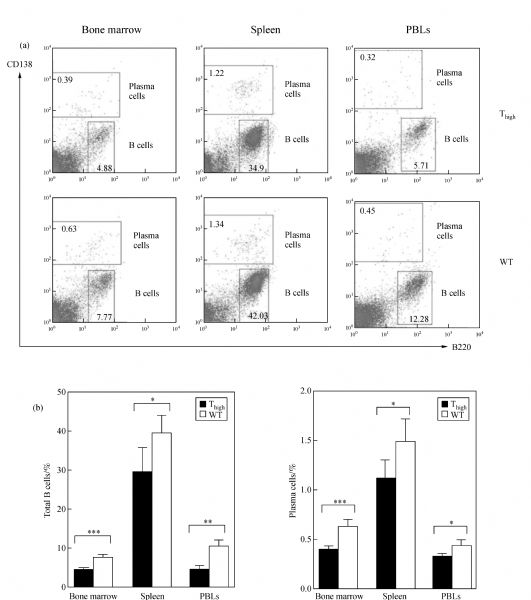
Abstract In this study, we introduced the bovine immunoglobulin μ heavy-chain gene (the orphaned gene on BTA11) into mouse germline cells. Bovine IgM was highly expressed in selected transgenic lines, and it largely inhibited rearrangements of the endogenous immunoglobulin heavy chain (IgH) genes in these lines. The forced expression of bovine IgM resulted in reduced numbers of pro- and pre-B cells but increased the number of immature B cells in the transgenic mice. Bovine IgM-expressing B cells can migrate from the bone marrow to the spleen, but most of the cells are arrested at the T1 transitional B cell stage, leading to a significantly lower number of T2 transitional and mature B cells in the spleen. Although the serum concentrations of endogenous IgM and IgG in the transgenic mice were significantly decreased, the IgA levels were slightly increased compared to the WT mice. The bovine IgM level in the serum was only one-tenth to one-fifth of that of endogenous mouse IgM, suggesting that most of the serum immunoglobulin were contributed by endogenous IgH gene-expressing B cells. These transgenic mice also exhibited a lower frequency of unique complementarity determining region 3 (CDR3) sequences in their VH repertoire and Vκ repertoire but exhibited an increased frequency of unique CDR3 in their Vλ repertoire. Compared to the WT mice, the transgenic mice had a significantly higher percentage of mouse IgM-expressing B cells that expressed λ chains. Finally, we showed that the transgenic mice were deficient in a specific antibody response to antigen stimulation.
|
| Keywords
bovine Ig μ heavy-chain
transgenic mice
B cell development
allelic exclusion
immune response
Ig repertoire
|
|
Corresponding Author(s):
Yaofeng ZHAO
|
|
Online First Date: 22 September 2014
Issue Date: 10 October 2014
|
|
| 1 |
Flajnik M F. Comparative analyses of immunoglobulin genes: surprises and portents. Nature Reviews-Immunology, 2002, 2(9): 688–698
https://doi.org/10.1038/nri889
pmid: 12209137
|
| 2 |
Flajnik M F, Kasahara M. Origin and evolution of the adaptive immune system: genetic events and selective pressures. Nature Reviews-Genetics, 2010, 11(1): 47–59
https://doi.org/10.1038/nrg2703
pmid: 19997068
|
| 3 |
Matthias P, Rolink A G. Transcriptional networks in developing and mature B cells. Nature Reviews-Immunology, 2005, 5(6): 497–508
https://doi.org/10.1038/nri1633
pmid: 15928681
|
| 4 |
Yancopoulos G D, Blackwell T K, Wood C, Thomas E, Boss M, Coffman R, Rosenberg N, Tonegawa S, Baltimore D. EMBO Journal, 1984, 3(6): 1209–1219
pmid: 6086308
|
| 5 |
ten Boekel E, Melchers F, Rolink A G. Precursor B cells showing H chain allelic inclusion display allelic exclusion at the level of pre-B cell receptor surface expression. Immunity, 1998, 8(2): 199–207
https://doi.org/10.1016/S1074-7613(00)80472-0
pmid: 9492001
|
| 6 |
Keyna U, Beck-Engeser G B, Jongstra J, Applequist S E, J?ck H M. Surrogate light chain-dependent selection of Ig heavy chain V regions. Journal of Immunology, 1995, 155(12): 5536–5542
pmid: 7499835
|
| 7 |
Kline G H, Hartwell L, Beck-Engeser G B, Keyna U, Zaharevitz S, Klinman N R, J?ck H M. Pre-B cell receptor-mediated selection of pre-B cells synthesizing functional mu heavy chains. Journal of Immunology, 1998, 161(4): 1608–1618
pmid: 9712022
|
| 8 |
Melchers F. The pre-B-cell receptor: selector of fitting immunoglobulin heavy chains for the B-cell repertoire. Nature Reviews-Immunology, 2005, 5(7): 578–584
https://doi.org/10.1038/nri1649
pmid: 15999097
|
| 9 |
Pernis B, Chiappino G, Kelus A S, Gell P G. Cellular localization of immunoglobulins with different allotypic specificities in rabbit lymphoid tissues. The Journal of Experimental Medicine, 1965, 122(5): 853–876
https://doi.org/10.1084/jem.122.5.853
pmid: 4159057
|
| 10 |
Weiler E. Differential activity of allelic gamma-globulin genes in antibody-producing cells. Proceedings of the National Academy of Sciences of the United States of America, 1965, 54(6): 1765–1772
https://doi.org/10.1073/pnas.54.6.1765
pmid: 4160302
|
| 11 |
Vettermann C, Schlissel M S. Allelic exclusion of immunoglobulin genes: models and mechanisms. Immunological Reviews, 2010, 237(1): 22–42
https://doi.org/10.1111/j.1600-065X.2010.00935.x
pmid: 20727027
|
| 12 |
Lutz J, Heideman M R, Roth E, van den Berk P, Müller W, Raman C, Wabl M, Jacobs H, J?ck H M. Pro-B cells sense productive immunoglobulin heavy chain rearrangement irrespective of polypeptide production. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(26): 10644–10649
https://doi.org/10.1073/pnas.1019224108
pmid: 21670279
|
| 13 |
Nussenzweig M C, Shaw A C, Sinn E, Danner D B, Holmes K L, Morse H C 3rd, Leder P. Allelic exclusion in transgenic mice that express the membrane form of immunoglobulin mu. Science, 1987, 236(4803): 816–819
https://doi.org/10.1126/science.3107126
pmid: 3107126
|
| 14 |
Rusconi S, K?hler G. Transmission and expression of a specific pair of rearranged immunoglobulin mu and kappa genes in a transgenic mouse line. Nature, 1985, 314(6009): 330–334
https://doi.org/10.1038/314330a0
pmid: 3920529
|
| 15 |
Weaver D, Costantini F, Imanishi-Kari T, Baltimore D. A transgenic immunoglobulin mu gene prevents rearrangement of endogenous genes. Cell, 1985, 42(1): 117–127
https://doi.org/10.1016/S0092-8674(85)80107-0
pmid: 3926323
|
| 16 |
Roth P E, Doglio L, Manz J T, Kim J Y, Lo D, Storb U. Immunoglobulin gamma 2b transgenes inhibit heavy chain gene rearrangement, but cannot promote B cell development. The Journal of Experimental Medicine, 1993, 178(6): 2007–2021
https://doi.org/10.1084/jem.178.6.2007
pmid: 8245779
|
| 17 |
Nussenzweig M C, Shaw A C, Sinn E, Campos-Torres J, Leder P. Allelic exclusion in transgenic mice carrying mutant human IgM genes. The Journal of Experimental Medicine, 1988, 167(6): 1969–1974
https://doi.org/10.1084/jem.167.6.1969
pmid: 3133444
|
| 18 |
Herzenberg L A, Stall A M, Braun J, Weaver D, Baltimore D, Herzenberg L A, Grosschedl R. Depletion of the predominant B-cell population in immunoglobulin mu heavy-chain transgenic mice. Nature, 1987, 329(6134): 71–73
https://doi.org/10.1038/329071a0
pmid: 3114639
|
| 19 |
Jasper P J, Rhee K J, Kalis S L, Sethupathi P, Yam P C, Zhai S K, Knight K L. B lymphocyte deficiency in IgH-transgenic rabbits. European Journal of Immunology, 2007, 37(8): 2290–2299
https://doi.org/10.1002/eji.200737191
pmid: 17628859
|
| 20 |
Zhao Y, Kacskovics I, Pan Q, Liberles D A, Geli J, Davis S K, Rabbani H, Hammarstrom L. Artiodactyl IgD: the missing link. Journal of Immunology, 2002, 169(8): 4408–4416
https://doi.org/10.4049/jimmunol.169.8.4408
pmid: 12370374
|
| 21 |
Tobin-Janzen T C, Womack J E. Comparative mapping of IGHG1, IGHM, FES, and FOS in domestic cattle. Immunogenetics, 1992, 36(3): 157–165
https://doi.org/10.1007/BF00661092
pmid: 1612649
|
| 22 |
Hayes H C, Petit E J. Mapping of the beta-lactoglobulin gene and of an immunoglobulin M heavy chain-like sequence to homoeologous cattle, sheep, and goat chromosomes. Mammalian Genome, 1993, 4(4): 207–210
https://doi.org/10.1007/BF00417564
pmid: 8499654
|
| 23 |
Zhao Y, Kacskovics I, Rabbani H, Hammarstrom L. Physical mapping of the bovine immunoglobulin heavy chain constant region gene locus. Journal of Biological Chemistry, 2003, 278(37): 35024–35032
https://doi.org/10.1074/jbc.M301337200
pmid: 12829708
|
| 24 |
Kuroiwa Y, Kasinathan P, Sathiyaseelan T, Jiao J A, Matsushita H, Sathiyaseelan J, Wu H, Mellquist J, Hammitt M, Koster J, Kamoda S, Tachibana K, Ishida I, Robl J M. Antigen-specific human polyclonal antibodies from hyperimmunized cattle. Nature Biotechnology, 2009, 27(2): 173–181
https://doi.org/10.1038/nbt.1521
pmid: 19151699
|
| 25 |
Eggen A, Gautier M, Billaut A, Petit E, Hayes H, Laurent P, Urban C, Pfister-Genskow M, Eilertsen K, Bishop M D. Construction and characterization of a bovine BAC library with four genome-equivalent coverage. Genetics Selection Evolution, 2001, 33(5): 543–548
https://doi.org/10.1186/1297-9686-33-5-543
pmid: 11712974
|
| 26 |
Hosseini A, Campbell G, Prorocic M, Aitken R. Duplicated copies of the bovine JH locus contribute to the Ig repertoire. International Immunology, 2004, 16(6): 843–852
https://doi.org/10.1093/intimm/dxh085
pmid: 15126417
|
| 27 |
Loder F, Mutschler B, Ray R J, Paige C J, Sideras P, Torres R, Lamers M C, Carsetti R. B cell development in the spleen takes place in discrete steps and is determined by the quality of B cell receptor-derived signals. The Journal of Experimental Medicine, 1999, 190(1): 75–89
https://doi.org/10.1084/jem.190.1.75
pmid: 10429672
|
| 28 |
Carsetti R, Rosado M M, Wardmann H. Peripheral development of B cells in mouse and man. Immunological Reviews, 2004, 197(1): 179–191
https://doi.org/10.1111/j.0105-2896.2004.0109.x
pmid: 14962195
|
| 29 |
Koti M, Kataeva G, Kaushik A K. Novel atypical nucleotide insertions specifically at VH-DH junction generate exceptionally long CDR3H in cattle antibodies. Molecular Immunology, 2010, 47(11–12): 2119–2128
https://doi.org/10.1016/j.molimm.2010.02.014
pmid: 20435350
|
| 30 |
Kaushik A K, Kehrli M E Jr, Kurtz A, Ng S, Koti M, Shojaei F, Saini S S. Somatic hypermutations and isotype restricted exceptionally long CDR3H contribute to antibody diversification in cattle. Veterinary Immunology and Immunopathology, 2009, 127(1–2): 106–113
https://doi.org/10.1016/j.vetimm.2008.09.024
pmid: 19012969
|
| 31 |
Shimizu A, Nussenzweig M C, Mizuta T R, Leder P, Honjo T. Immunoglobulin double-isotype expression by trans-mRNA in a human immunoglobulin transgenic mouse. Proceedings of the National Academy of Sciences of the United States of America, 1989, 86(20): 8020–8023
https://doi.org/10.1073/pnas.86.20.8020
pmid: 2510157
|
| 32 |
Shimizu A, Nussenzweig M C, Han H, Sanchez M, Honjo T. Trans-splicing as a possible molecular mechanism for the multiple isotype expression of the immunoglobulin gene. The Journal of Experimental Medicine, 1991, 173(6): 1385–1393
https://doi.org/10.1084/jem.173.6.1385
pmid: 1903429
|
| 33 |
Ekman A, Niku M, Liljavirta J, Iivanainen A. Bos taurus genome sequence reveals the assortment of immunoglobulin and surrogate light chain genes in domestic cattle. BMC Immunology, 2009, 10(1): 22–32
https://doi.org/10.1186/1471-2172-10-22
pmid: 19405939
|
| 34 |
Stall A M, Kroese F G, Gadus F T, Sieckmann D G, Herzenberg L A, Herzenberg L A. Rearrangement and expression of endogenous immunoglobulin genes occur in many murine B cells expressing transgenic membrane IgM. Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(10): 3546–3550
https://doi.org/10.1073/pnas.85.10.3546
pmid: 3130629
|
| 35 |
Forni L. Extensive splenic B cell activation in IgM-transgenic mice. European Journal of Immunology, 1990, 20(5): 983–989
https://doi.org/10.1002/eji.1830200506
pmid: 2358019
|
| 36 |
Barreto V, Cumano A. Frequency and characterization of phenotypic Ig heavy chain allelically included IgM-expressing B cells in mice. Journal of Immunology, 2000, 164(2): 893–899
https://doi.org/10.4049/jimmunol.164.2.893
pmid: 10623837
|
| 37 |
Roes J, Rajewsky K. Immunoglobulin D (IgD)-deficient mice reveal an auxiliary receptor function for IgD in antigen-mediated recruitment of B cells. The Journal of Experimental Medicine, 1993, 177(1): 45–55
https://doi.org/10.1084/jem.177.1.45
pmid: 8418208
|
| 38 |
Nitschke L, Kosco M H, K?hler G, Lamers M C. Immunoglobulin D-deficient mice can mount normal immune responses to thymus-independent and -dependent antigens. Proceedings of the National Academy of Sciences of the United States of America, 1993, 90(5): 1887–1891
https://doi.org/10.1073/pnas.90.5.1887
pmid: 8446604
|
| 39 |
Knott J, Bona C, Kaushik A. The primary antibody repertoire of κ-deficient mice is characterized by non-stochastic Vλ1 + VH gene family pairings and a higher degree of self-reactivity. Scandinavian Journal of Immunology, 1998, 48(1): 65–72
https://doi.org/10.1046/j.1365-3083.1998.00369.x
pmid: 9714412
|
| 40 |
Khan W N. B cell receptor and BAFF receptor signaling regulation of B cell homeostasis. Journal of Immunology, 2009, 183(6): 3561–3567
https://doi.org/10.4049/jimmunol.0800933
pmid: 19726767
|
| 41 |
Macpherson A J, Lamarre A, McCoy K, Harriman G R, Odermatt B, Dougan G, Hengartner H, Zinkernagel R M. IgA production without mu or delta chain expression in developing B cells. Nature Immunology, 2001, 2(7): 625–631
https://doi.org/10.1038/89775
pmid: 11429547
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|