|
|
|
Plant stem cells and their regulations in shoot apical meristems |
Chun-Ming LIU( ), Yuxin HU() ), Yuxin HU() |
| Center for Signal Transduction & Metabolomics, Key Laboratory of Photosynthesis and Environmental Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China |
|
|
|
|
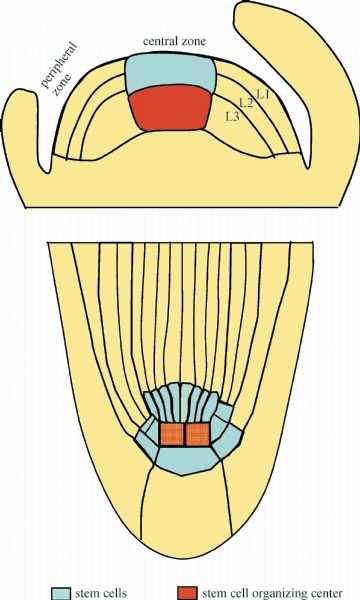
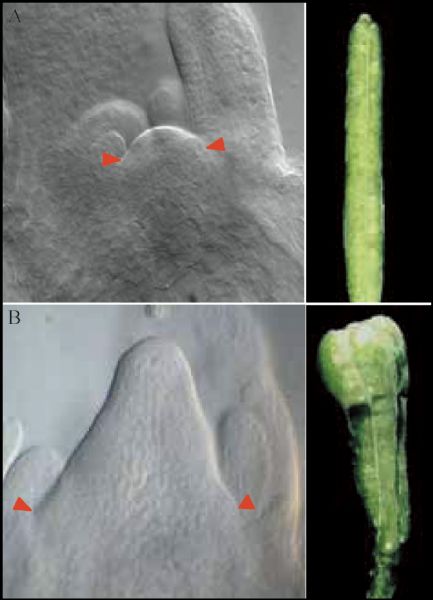
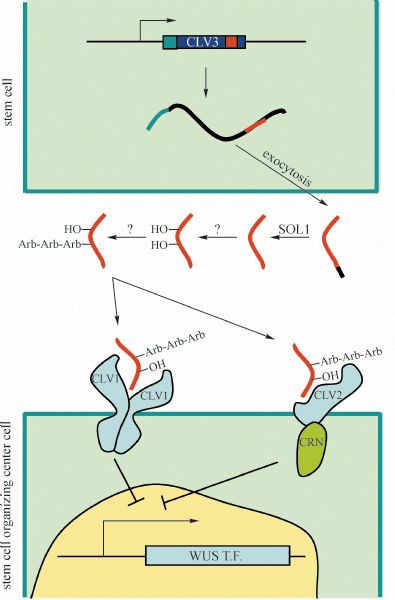
Abstract Stem cells in plants, established during embryogenesis, are located in the centers of the shoot apical meristem (SAM) and the root apical meristem (RAM). Stem cells in SAM have a capacity to renew themselves and to produce new organs and tissues indefinitely. Although fully differentiated organs such as leaves do not contain stem cells, cells in such organs do have the capacity to re-establish new stem cells, especially under the induction of phytohormones in vitro. Cytokinin and auxin are critical in creating position signals in the SAM to maintain the stem cell organizing center and to position the new organ primordia, respectively. This review addresses the distinct features of plant stem cells and focuses on how stem cell renewal and differentiation are regulated in SAMs.
|
| Keywords
plant
stem cell
shoot apical meristem
root apical meristem
|
|
Corresponding Author(s):
LIU Chun-Ming,Email:cmliu@ibcas.ac.cn; HU Yuxin,Email:huyuxin@ibcas.ac.cn
|
|
Issue Date: 01 October 2010
|
|
| 1 |
Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh Y S, Amasino R, Scheres B (2004). The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell , 119(1): 109–120
|
| 2 |
Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J (2003). Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell , 115(5): 591–602
|
| 3 |
Brand U, Fletcher J C, Hobe M, Meyerowitz E M, Simon R (2000). Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science , 289(5479): 617–619
doi: 10.1126/science.289.5479.617
|
| 4 |
Casamitjana-Martínez E, Hofhuis H F, Xu J, Liu C M, Heidstra R, Scheres B (2003). Root-specific CLE19 overexpression and the sol/2 suppressors implicate a CLV-like pathway in the control of Arabidopsis root meristem maintenance. Curr Biol , 13(16): 1435–1441
doi: 10.1016/S0960-9822(03)00533-5
|
| 5 |
Chu H, Qian Q, Liang W, Yin C, Tan H, Yao X, Yuan Z, Yang J, Huang H, Luo D, Ma H, Zhang D (2006). The FLORAL ORGAN NUMBER4 organ number4 gene encoding a putative ortholog of Arabidopsis CLAVATA3 regulates apical meristem size in rice. Plant Physiol , 142(3): 1039–1052
doi: 10.1104/pp.106.086736
|
| 6 |
Clark S E, Williams R W, Meyerowitz E M (1997). The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell , 89(4): 575–585
|
| 7 |
Cock J M, McCormick S (2001). A large family of genes that share homology with CLAVATA3. Plant Physiol , 126(3): 939–942
doi: 10.1104/pp.126.3.939
|
| 8 |
Fiers M, Golemiec E, van der Schors R, van der Geest L, Li K W, Stiekema W J, Liu C M (2006). The CLAVATA3/ESR motif of CLAVATA3 is functionally independent from the nonconserved flanking sequences. Plant Physiol , 141(4): 1284–1292
doi: 10.1104/pp.106.080671
|
| 9 |
Fiers M, Golemiec E, Xu J, van der Geest L, Heidstra R, Stiekema W, Liu C M (2005). The 14-amino acid CLV3, CLE19, and CLE40 peptides trigger consumption of the root meristem in Arabidopsis through a CLAVATA2-dependent pathway. Plant Cell , 17(9): 2542–2553
doi: 10.1105/tpc.105.034009
|
| 10 |
Fiers M, Ku K L, Liu C M (2007). CLE peptide ligands and their roles in establishing meristems. Curr Opin Plant Biol , 10(1): 39–43
doi: 10.1016/j.pbi.2006.11.003
|
| 11 |
Fletcher J C, Brand U, Running M P, Simon R, Meyerowitz E M (1999). Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science , 283(5409): 1911–1914
doi: 10.1126/science.283.5409.1911
|
| 12 |
Geier F, Lohmann J U, Gerstung M, Maier A T, Timmer J, Fleck C (2008). A quantitative and dynamic model for plant stem cell regulation. PLoS One , 3(10): e3553
doi: 10.1371/journal.pone.0003553
|
| 13 |
Gordon S P, Heisler M G, Reddy G V, Ohno C, Das P, Meyerowitz E M (2007). Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development , 134(19): 3539–3548
doi: 10.1242/dev.010298
|
| 14 |
Gross-Hardt R, Laux T (2003). Stem cell regulation in the shoot meristem. J Cell Sci , 116(Pt 9): 1659–1666
doi: 10.1242/jcs.00406
|
| 15 |
Hass C, Lohrmann J, Albrecht V, Sweere U, Hummel F, Yoo S D, Hwang I, Zhu T, Sch?fer E, Kudla J, Harter K (2004). The response regulator 2 mediates ethylene signalling and hormone signal integration in Arabidopsis. EMBO J , 23(16): 3290–3302
doi: 10.1038/sj.emboj.7600337
|
| 16 |
Heisler M G, Ohno C, Das P, Sieber P, Reddy G V, Long J A, Meyerowitz E M (2005). Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr Biol , 15(21): 1899–1911
doi: 10.1016/j.cub.2005.09.052
|
| 17 |
Hobe M, Müller R, Grünewald M, Brand U, Simon R (2003). Loss of CLE40, a protein functionally equivalent to the stem cell restricting signal CLV3, enhances root waving in Arabidopsis. Dev Genes Evol , 213(8): 371–381
doi: 10.1007/s00427-003-0329-5
|
| 18 |
Hwang I, Sheen J (2001). Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature , 413(6854): 383–389
doi: 10.1038/35096500
|
| 19 |
Jeong S, Trotochaud A E, Clark S E (1999). The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell , 11(10): 1925–1934
|
| 20 |
J?nsson H, Heisler M, Reddy G V, Agrawal V, Gor V, Shapiro B E, Mjolsness E, Meyerowitz E M (2005). Modeling the organization of the WUSCHEL expression domain in the shoot apical meristem. Bioinformatics , 21(Suppl 1): i232–i240
doi: 10.1093/bioinformatics/bti1036
|
| 21 |
Kondo T, Sawa S, Kinoshita A, Mizuno S, Kakimoto T, Fukuda H, Sakagami Y (2006). A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science , 313(5788): 845–848
doi: 10.1126/science.1128439
|
| 22 |
Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, Nagato Y, Sakakibara H, Kyozuka J (2007). Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature , 445(7128): 652–655
doi: 10.1038/nature05504
|
| 23 |
Laux T, Mayer K F, Berger J, Jürgens G (1996). The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development , 122(1): 87–96
|
| 24 |
Leibfried A, To J P, Busch W, Stehling S, Kehle A, Demar M, Kieber J J, Lohmann J U (2005). WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature , 438(7071): 1172–1175
doi: 10.1038/nature04270
|
| 25 |
Lenhard M, Bohnert A, Jürgens G, Laux T (2001). Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell , 105(6): 805–814
|
| 26 |
Lenhard M, Laux T (2003). Stem cell homeostasis in the Arabidopsis shoot meristem is regulated by intercellular movement of CLAVATA3 and its sequestration by CLAVATA1. Development , 130(14): 3163–3173
doi: 10.1242/dev.00525
|
| 27 |
Leyser H M O, Furner I J (1992). Characterisation of three shoot apical meristem mutants of Arabidopsis thaliana. Development , 116: 397–403
|
| 28 |
Lindsay D L, Sawhney V K, Bonham-Smith P C (2006). Cytokinin-induced changes in CLAVATA1 and WUSCHEL expression temporally coincide with altered floral development in Arabidopsis. Plant Sci , 170: 1111–1117
doi: 10.1016/j.plantsci.2006.01.015
|
| 29 |
Liu C M, Xu Z, Chua N H (1993). Auxin Polar transport is essential for the establishment of bilateral symmetry during early plant embryogenesis. Plant Cell , 5(6): 621–630
|
| 30 |
Mayer K F, Schoof H, Haecker A, Lenhard M, Jürgens G, Laux T (1998). Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell , 95(6): 805–815
|
| 31 |
Müller B, Sheen J (2008). Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature , 453(7198): 1094–1097
doi: 10.1038/nature06943
|
| 32 |
Müller R, Bleckmann A, Simon R (2008). The receptor kinase CORYNE of Arabidopsis transmits the stem cell-limiting signal CLAVATA3 independently of CLAVATA1. Plant Cell , 20(4): 934–946
doi: 10.1105/tpc.107.057547
|
| 33 |
Nishimura C, Ohashi Y, Sato S, Kato T, Tabata S, Ueguchi C (2004). Histidine kinase homologs that act as cytokinin receptors possess overlapping functions in the regulation of shoot and root growth in Arabidopsis. Plant Cell , 16(6):1365–1377
doi: 10.1105/tpc.021477
|
| 34 |
Ogawa M, Shinohara H, Sakagami Y, Matsubayashi Y (2008). Arabidopsis CLV3 peptide directly binds CLV1 ectodomain. Science , 319(5861): 294
doi: 10.1126/science.1150083
|
| 35 |
Ohyama K, Shinohara H, Ogawa-Ohnishi M, Matsubayashi Y (2009). A glycopeptide regulating stem cell fate in Arabidopsis thaliana. Nat Chem Biol , 5(8): 578–580
doi: 10.1038/nchembio.182
|
| 36 |
Philipson W R (1954). Organization of the shoot apex in dicotyledons. Phytomorphogy , 4: 70–75
|
| 37 |
Reinhardt D, Frenz M, Mandel T, Kuhlemeier C (2003). Microsurgical and laser ablation analysis of interactions between the zones and layers of the tomato shoot apical meristem. Development , 130(17): 4073–4083
doi: 10.1242/dev.00596
|
| 38 |
Reinhardt D, Mandel T, Kuhlemeier C (2000). Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell , 12(4): 507–518
|
| 39 |
Riefler M, Novak O, Strnad M, Schmülling T (2006). Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell , 18(1): 40–54
doi: 10.1105/tpc.105.037796
|
| 40 |
Sabatini S, Heidstra R, Wildwater M, Scheres B (2003). SCARECROW is involved in positioning the stem cell niche in the Arabidopsis root meristem. Genes Dev , 17(3): 354–358
doi: 10.1101/gad.252503
|
| 41 |
Sakai H, Aoyama T, Oka A (2000). Arabidopsis ARR1 and ARR2 response regulators operate as transcriptional activators. Plant J , 24(6): 703–711
doi: 10.1046/j.1365-313x.2000.00909.x
|
| 42 |
Scheres B (2007). Stem-cell niches: nursery rhymes across kingdoms. Nat Rev Mol Cell Biol , 8(5): 345–354
doi: 10.1038/nrm2164
|
| 43 |
Schoof H, Lenhard M, Haecker A, Mayer K F X, Jürgens G, Laux T (2000). The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell , 100(6): 635–644
|
| 44 |
Singh M B, Bhalla P L (2006). Plant stem cells carve their own niche. Trends Plant Sci , 11(5): 241–246
doi: 10.1016/j.tplants.2006.03.004
|
| 45 |
Skoog F, Miller C O (1957). Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp Soc Exp Biol , 54(11): 118–130
|
| 46 |
Steeves T A, Sussex I M (1989). Patterns in plant development. 2nd ed. Cambridge: Cambridge University Press
doi: 10.1017/CBO9780511626227
|
| 47 |
Su Y H, Zhao X Y, Liu Y B, Zhang C L, O’Neill S D, Zhang X S (2009). Auxin-induced WUS expression is essential for embryonic stem cell renewal during somatic embryogenesis in Arabidopsis. Plant J , 59(3): 448–460
doi: 10.1111/j.1365-313X.2009.03880.x
|
| 48 |
Sussex I M (1955). Morphologenesis in Solanum tuberosum L.: Apical structure and developmental pattern of the juvenile shoot. Phytomorphology , 5: 253–273
|
| 49 |
Swarup K, Benková E, Swarup R, Casimiro I, Péret B, Yang Y, Parry G, Nielsen E, De Smet I, Vanneste S, Levesque M P, Carrier D, James N, Calvo V, Ljung K, Kramer E, Roberts R, Graham N, Marillonnet S, Patel K, Jones J D, Taylor C G, Schachtman D P, May S, Sandberg G, Benfey P, Friml J, Kerr I, Beeckman T, Laplaze L, Bennett M J (2008). The auxin influx carrier LAX3 promotes lateral root emergence. Nat Cell Biol , 10(8): 946–954
doi: 10.1038/ncb1754
|
| 50 |
To J P, Haberer G, Ferreira F J, Deruère J, Mason M G, Schaller G E, Alonso J M, Ecker J R, Kieber J J (2004). Type-A Arabidopsis response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell , 16(3): 658–671
doi: 10.1105/tpc.018978
|
| 51 |
Trotochaud A E, Hao T, Wu G, Yang Z, Clark S E (1999). The CLAVATA1 receptor-like kinase requires CLAVATA3 for its assembly into a signaling complex that includes KAPP and a Rho-related protein. Plant Cell , 11(3): 393–406
|
| 52 |
van den Berg C, Willemsen V, Hage W, Weisbeek P, Scheres B (1995). Cell fate in the Arabidopsis root meristem determined by directional signalling. Nature , 378(6552): 62–65
doi: 10.1038/378062a0
|
| 53 |
van den Berg C, Willemsen V, Hendriks G, Weisbeek P, Scheres B (1997). Short-range control of cell differentiation in the Arabidopsis root meristem. Nature , 390(6657): 287–289
doi: 10.1038/36856
|
| 54 |
Zhu Y F, Wang Y, Li R, Song X, Wang Q, Huang S, Jin J B, Liu C M, Lin J X (2010). Analysis of interactions among the CLAVATA3 receptors reveals a direct interaction between CLAVATA2 and CORYNE in Arabidopsis. Plant J , 61(2): 223–233
doi: 10.1111/j.1365-313X.2009.04049.x
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|